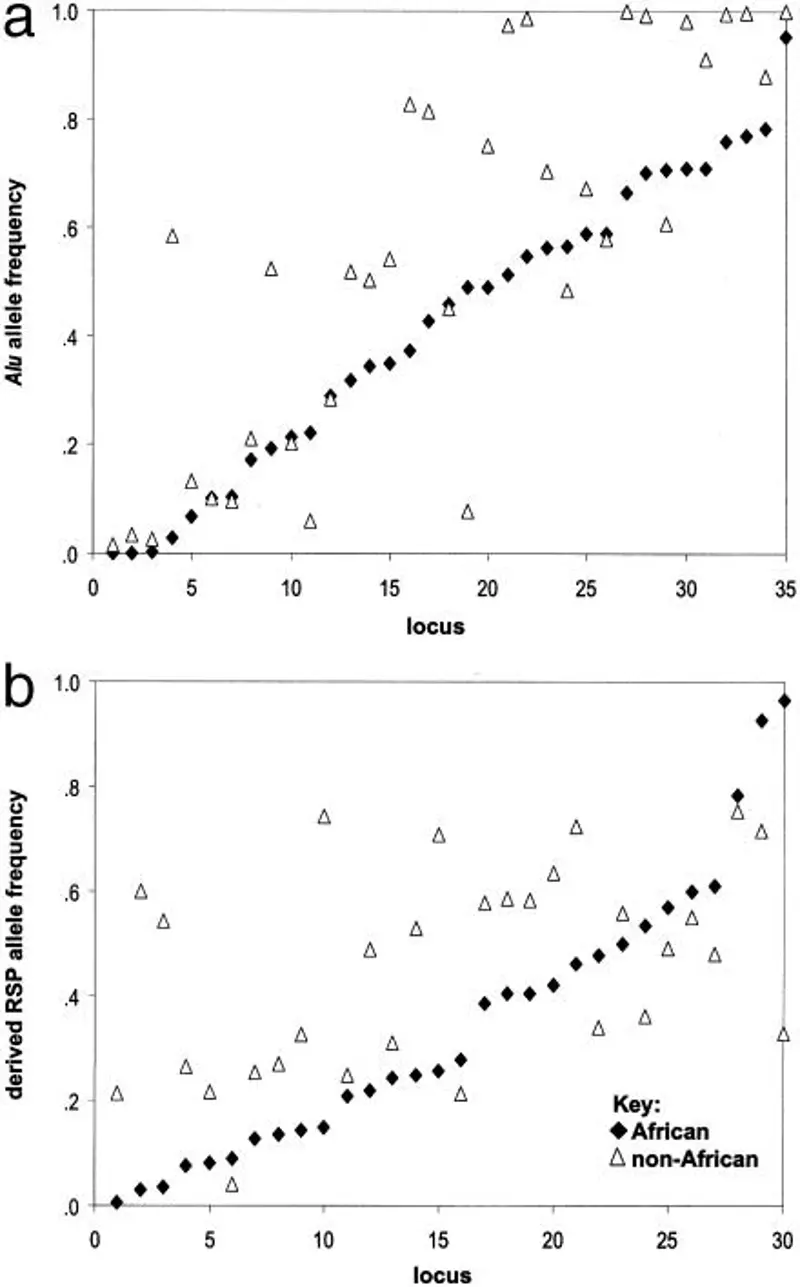
Obecność archaicznych form wczesnych homo w populacjach afrykańskich. Drzewa są zakorzenione w populacjach afrykańskich (np. Mbuti i Zair Pygmy), które są genetycznie najbliższe populacji przodków. Spośród 35 polimorficznych insercji Alu, 33 występują we wszystkich grupach kontynentalnych. Insercje Ya5NBC54 i Ya5NBC135 nie zostały znalezione w populacjach afrykańskich. We wszystkich subpopulacjach poza Afryką zaobserwowano wyższy poziom homozygotyczności Alu, uśredniony dla 35 loci Alu, niż we wszystkich populacjach w Afryce. Można dostrzec dwa wzorce: (1) populacje afrykańskie wykazują ogólną tendencję do niższych częstości insercji Alu oraz (2) loci, które mają niską (<.3) częstość u osób niebędących Afrykanami, mają również niskie częstości u Afrykanów, podczas gdy loci o wyższych częstościach u osób niebędących Afrykanami mają tendencję do niższych częstości u Afrykanów. Te wzorce zaobserwowano również, gdy grupy kontynentalne podzielono na subpopulacje. Subpopulacje afrykańskie wykazywały tendencje do niższych częstości insercji Alu, podczas gdy najwyższe częstości insercji stwierdzono w populacjach Azji Południowo-Wschodniej. Średnie częstości insercji Alu dla głównych grup populacji są następujące: Afrykanie, 0,4226: Azjaci, 0,5690: Europejczycy, 0,5604: i Hindusi, 0,5453. Rozkłady częstości alleli w czterech głównych grupach populacji istotnie różnią się między Afrykanami i Azjatami (p <.001, na podstawie testu znaków Wilcoxona), Afrykanami i Europejczykami (p <.003) oraz Afrykanami i Hindusami (p <.002), ale nie różnią się istotnie między grupami nieafrykańskimi. Do utworzenia populacji przodków wykorzystano pochodną częstość allelu RSP (0) w każdym locus. Populacja przodków w sieci RSP jest bardzo zbliżona do klastra afrykańskiego. Populacje afrykańskie odgałęziają się jako pierwsze, a wsparcie bootstrapowe dla klastra nieafrykańskiego jest bardzo wysokie. Najnowsze badania wykorzystujące niezależne zestawy STRP i markerów biallelicznych na chromosomie Y sugerują, że najwcześniejsze odgałęzienia męskiego/męskiego założyciela/założycieli naszego gatunku można prześledzić do przodków afrykańskiej populacji San (Forster i in. 2000). W sieciach Alu i RSP osobniki San badane w tej analizie znajdują się w pobliżu populacji przodków. Szacunki heterozygotyczności i homozygotyczności Alu dla grup kontynentalnych i ich subpopulacji są zgodne z rozkładami częstości Alu (tabela 1a). Populacje afrykańskie charakteryzują się większą ogólną różnorodnością genu Alu (heterozygotycznością) niż inne populacje. Odkrycie to jest zgodne z wynikami innych systemów markerowych, w tym genomu mitochondrialnego, autosomalnych STRP i chromosomu Y (Deka i in. 1995: Jorde i in. 2000). Obserwowana homozygotyczność dla insertu Alu (stan pochodny) jest istotnie wyższa (p < 0,05) we wszystkich grupach nieafrykańskich niż u Afrykanów Subsaharyjskich. Wszystkie subpopulacje poza Afryką charakteryzują się wyższymi obserwowanymi poziomami homozygotyczności Alu, uśrednionymi dla 35 loci Alu, niż wszystkie populacje w Afryce. Porównania głównych grup populacji i ich subpopulacji, z wykorzystaniem zakorzenionych loci RSP, wykazują podobną tendencję, poza Afryką, w kierunku wzrostu częstości i homozygotyczności alleli pochodnych (tabela 1b). Pomimo wysokiego poziomu heterozygotyczności w Europie, spowodowanego błędem w ocenie (Bowcock i in. 1991: Rogers i Jorde 1996), obserwowany poziom homozygotyczności RSP alleli pochodnych, we wszystkich loci, jest najniższy w Afryce i jej subpopulacjach, wyższy w Europie i najwyższy w populacjach Azji Południowo-Wschodniej. Sieci łączące się z sąsiadami oparte na insercjach Alu lub RSP mają swoje korzenie w Afryce i ukazują populacje afrykańskie jako odrębne od innych populacji, z wysokim wsparciem statystycznym. Korelacje między odległościami genetycznymi opartymi na Alu i jądrowych RSP, krótkich polimorfizmach powtórzeń tandemowych i mtDNA u tych samych osobników są wysokie i istotne. Dla 35 loci, różnorodność genu Alu i różnorodność przypisywana podziałowi populacji jest najwyższa w Afryce, ale jest niższa i podobna w Europie i Azji. Rozmieszczenie alleli przodków jest zgodne z pochodzeniem wczesnych populacji człowieka współczesnego w Afryce Subsaharyjskiej, izolacją i zachowaniem alleli przodków w Afryce oraz ekspansją z Afryki do Eurazji. Ekspansja ta charakteryzuje się rosnącą częstością insercji Alu i pochodnymi alleli RSP przy zmniejszonej różnorodności genetycznej w populacjach nieafrykańskich.
Komentarze
Napisz pierwszy komentarz!