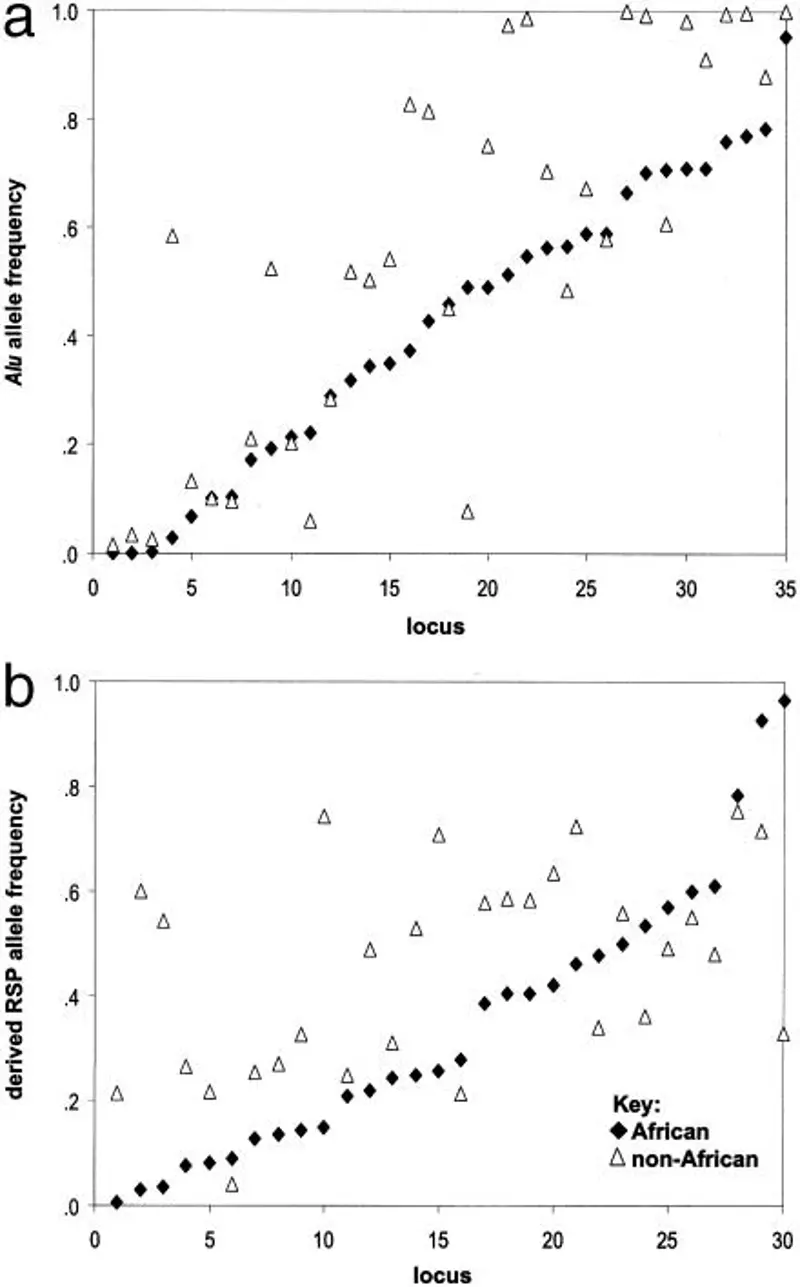
Presence of archaic forms of early Homo in African populations. The trees are rooted in African populations (e.g., Mbuti and Zaire Pygmy), which are genetically closest to the ancestral population. Of the 35 polymorphic Alu insertions, 33 are found in all continental groups. The Ya5NBC54 and Ya5NBC135 insertions were not found in African populations. A higher level of Alu homozygosity, averaged over 35 Alu loci, was observed in all subpopulations outside Africa than in all populations in Africa. Two patterns can be discerned: (1) African populations show an overall trend toward lower Alu insertion frequencies, and (2) loci that have low (<.3) frequencies in non-Africans also have low frequencies in Africans, while loci with higher frequencies in non-Africans tend to have lower frequencies in Africans. These patterns were also observed when continental groups were divided into subpopulations. African subpopulations tended to have lower Alu insertion frequencies, while the highest insertion frequencies were found in Southeast Asian populations. The average Alu insertion frequencies for the major population groups are as follows: Africans, 0.4226: Asians, 0.5690: Europeans, 0.5604: and Indians, 0.5453. The allele frequency distributions for the four major population groups are significantly different between Africans and Asians (p <.001, based on Wilcoxon sign test), Africans and Europeans (p <.003), and Africans and Indians (p <.002), but not significantly different between non-African groups. The derived RSP allele frequency (0) at each locus was used to create the ancestral population. The ancestral population in the RSP network is very similar to the African cluster. The African populations branch out first, and bootstrap support for the non-African cluster is very high. Recent studies using independent sets of STRPs and biallelic markers on the Y chromosome suggest that the earliest branching male/male founder(s) of our species can be traced to the ancestors of the African San population (Forster et al. 2000). In the Alu and RSP networks, the San individuals studied in this analysis are close to the ancestral population. Estimates of Alu heterozygosity and homozygosity for continental groups and their subpopulations are consistent with Alu frequency distributions (Table 1a). African populations have greater overall Alu gene diversity (heterozygosity) than other populations. This finding is consistent with results from other marker systems, including the mitochondrial genome, autosomal STRPs and the Y chromosome (Deka et al. 1995: Jorde et al. 2000). The observed homozygosity for the Alu insertion (derived state) is significantly higher (p < 0.05) in all non-African groups than in sub-Saharan Africans. All non-African subpopulations have higher observed levels of Alu homozygosity, averaged over 35 Alu loci, than all African populations. Comparisons of major population groups and their subpopulations, using rooted RSP loci, show a similar trend, outside Africa, toward increased frequency and homozygosity of derived alleles (Table 1b). Despite the high level of heterozygosity in Europe, due to a bias (Bowcock et al. 1991: Rogers and Jorde 1996), the observed level of RSP homozygosity of derived alleles, at all loci, is lowest in Africa and its subpopulations, higher in Europe and highest in Southeast Asian populations. Neighbor-joining networks based on Alu or RSP insertions have their roots in Africa and show African populations as distinct from other populations, with high statistical support. Correlations between genetic distances based on Alu and nuclear RSPs, short tandem repeat polymorphisms and mtDNA in the same individuals are high and significant. For 35 loci, the diversity of the Alu gene and the diversity attributed to population sharing is highest in Africa, but is lower and similar in Europe and Asia. The distribution of ancestral alleles is consistent with the origin of early modern human populations in sub-Saharan Africa, the isolation and conservation of ancestral alleles in Africa, and the expansion from Africa to Eurasia. This expansion is characterized by an increasing frequency of Alu insertions and derived RSP alleles with reduced genetic diversity in non-African populations.
Comments
Be the first to comment!